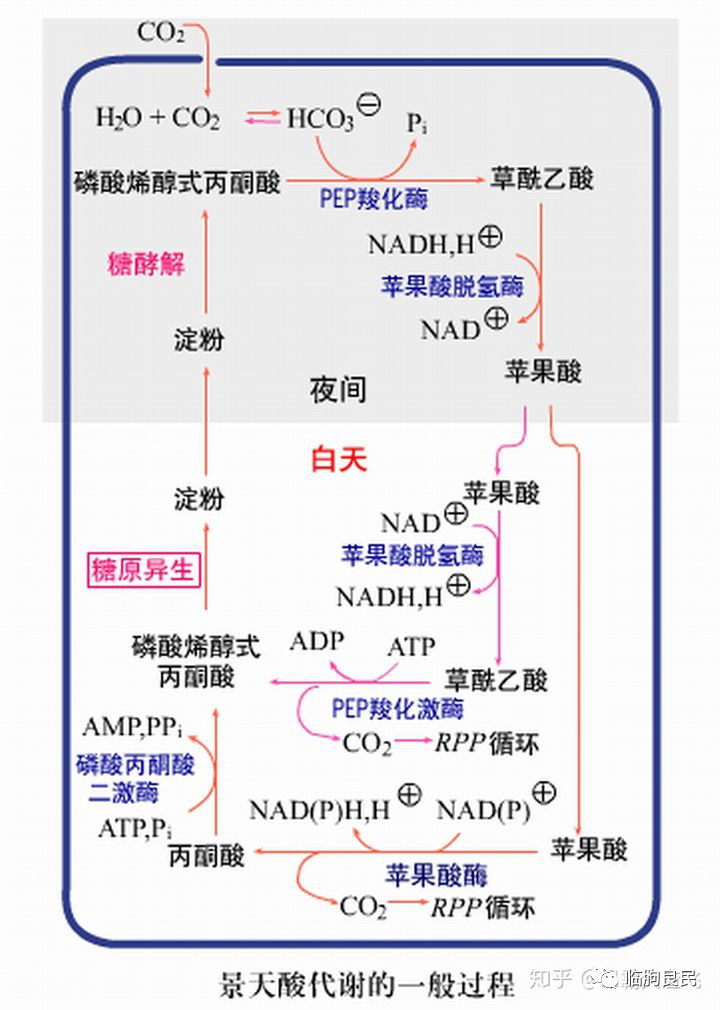
在自然环境条件中,光强度和温度的变化紧密交织在一起。在所有的细胞器官中,只有叶绿体同时对这两种变化展示出强烈的反应。越来越多的证据表明叶绿体代谢的变化对于植物适应环境至关重要,面对气候变化的严峻挑战,这些知识将辅助我们进行抗逆型的育种工作。

2022年01月07日,德国凯泽斯劳滕大学的H. EkkehardNeuhaus团队在Trends in Plant Science杂志在线发表题为“Chloroplasts are key players to cope with light and temperature stress”的综述性论文。改论文回顾了对于叶绿体的研究,并强调了叶绿体在植物应对光照与温度胁迫中的关键作用。
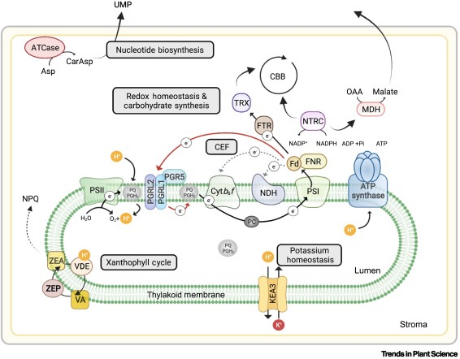
图1. 面对高光或低温胁迫,类囊体或基质中的快速代谢反应ATCase, aspartate transcarbamoylase; CBC, Calvin-Benson cycle; CEF, cyclic electron flow; CTP, cytidine triphosphate; Fd, ferredoxin; KEA3, potassium exchanger3; LEF, linear electron flow; NPQ, non-photochemical quenching; NTRC, NADP-dependent thioredoxin reductase C; OAA, oxaloacetate; pmf, proton motive force; PSI, photosystem I; TRX, thioredoxin; UTP, uridine triphosphate; VA, violaxanthin; VDE, violaxanthin de-epoxidase; ZEP, zeaxanthin.
为了平衡能量供应与能量消耗,叶绿体必须进行实时调整以适应外界的环境变化(图1)。LEF或CEF中的电子流会影响基质中的ATP:NADPH比,并影响穿过类囊体薄膜的pmf。pmf穿过类囊体薄膜这一过程也受到由NPQ介导的光能耗散的影响。PGR5是CEF的核心调节器,它与PGRL1以及PGRL2紧密互作,并且可以在部分没有PGRL1的情况下发挥作用。当处于弱光条件下,NPQ会急剧降低,此时由KEA3介导的钾离子转运具有非常关键的作用,KEA3介导的钾离子向类囊体内腔的运输可以抑制高量的pmf。大量的pmf通过类囊体薄膜会激活VA的积累,这个过程也会影响到NPQ。提高VDE与ZEP活性的突变体展现出产量的提升。NTRC负责基质内的还原平衡,这个过程利用NADPH作为电子的供体,并涉及2Cys-peroxiredoxins,CBC,NADP-MDH。NADP-MDH能够通过苹果酸/OAA的运输诱导等价的还原物发生。ATCase定位于寒冷条件下的类囊体薄膜内表面,因此可能将UTP和CTP的前体天门冬氨酰转移到核苷酸生物合成的胞质部分。这使核苷酸生物合成在低温下保持较高水平,与冷诱导的核糖体细胞积累一致,从而在这种不利条件下保持较高的mRNA翻译效率。
图2.适应高光和低温所需的运输过程的修饰ATCase, aspartate transcarbamoylase; OPDA, oxophytodieonic acid; RBL, rhomboid-like proteases; ROS, reactive oxygen species; TIC, inner translocon; TOC, outer translocon.
为了平衡外界的环境挑战,叶绿体作为中枢执行和整合由非生物环境变化引起的广泛生理反应。因为叶绿体与细胞的其余部分紧密结合,这个细胞器通常作为信使分子的发射器和接收器(图2)。由AtPHT4;4介导的抗坏血酸输入是高耐光性的关键。一些抗坏血酸的前体蛋白通过STY型蛋白激酶进行磷酸化以获得导入能力,这些蛋白激酶在基因表达和代谢物相互作用水平上受到调节。茉莉酸对启动获得耐寒性所需的遗传程序至关重要,茉莉酸盐前体OPDA依赖载体JASSY进行运输并通过叶绿体外膜。为了保持基质中保护性麦芽糖的高水平,在寒冷条件开始后,唯一的麦芽糖输出物(MEX)1的丰度显著下调。而膜结合蛋白酶如FTSH11或菱形蛋白酶是否有助于这种现象尚不清楚。
本文回顾的调控过程为叶绿体在适应光强度和温度的变化中的核心作用提供了有力的证据,但仍然有许多环节的研究是缺失的,在这里文章作者总结了几个仍待解决的问题:
1. 哪些蛋白质直接参与CEF,这一过程是如何调节的?PGR5是如何在不具备典型电子转运体特征的情况下促进电子转移的?
2. 为什么PGR5单独存在时比只缺少PGR5或同时缺少PGR1、PGR2、PGR5三种蛋白质更有害于植物生长?
3. 如何调节细胞氧化还原景观,以整合类囊体能量转导、代谢和基因表达,使其在波动环境中正常的生长?
4. 为什么植物核苷酸生物合成的一部分位于基质中,而所有其他生物体的这一途径通常位于细胞质中?
5. 为什么植物ATCase的结构与其他生物体中相应的同系物如此不同?
6. 哪种转运蛋白负责OPDA跨内膜的输出?
7. 在寒冷环境中,能快速有效地降解囊膜载体的蛋白酶分子特质是什么?
8. 蛋白质如何在低温下导入叶绿体?
原文链接:
限 时 特 惠: 本站每日持续更新海量各大内部创业教程,一年会员只需98元,全站资源免费下载 点击查看详情
站 长 微 信: lzxmw777